Everybody seems to be buzzing about this study on Atapuerca (El Portalón) site's Chalcolithic and Bronze Age genomes and I do think it is indeed worth taking a good look.
Torsten Günther, Cristina Valdiosera et al., Ancient genomes link early farmers from Atapuerca in Spain to modern-day Basques. PNAS 2015. Open access → LINK [doi: 10.1073/pnas.1509851112]
Abstract
The consequences of the Neolithic transition in Europe—one of the most important cultural changes in human prehistory—is a subject of great interest. However, its effect on prehistoric and modern-day people in Iberia, the westernmost frontier of the European continent, remains unresolved. We present, to our knowledge, the first genome-wide sequence data from eight human remains, dated to between 5,500 and 3,500 years before present, excavated in the El Portalón cave at Sierra de Atapuerca, Spain. We show that these individuals emerged from the same ancestral gene pool as early farmers in other parts of Europe, suggesting that migration was the dominant mode of transferring farming practices throughout western Eurasia. In contrast to central and northern early European farmers, the Chalcolithic El Portalón individuals additionally mixed with local southwestern hunter–gatherers. The proportion of hunter–gatherer-related admixture into early farmers also increased over the course of two millennia. The Chalcolithic El Portalón individuals showed greatest genetic affinity to modern-day Basques, who have long been considered linguistic and genetic isolates linked to the Mesolithic whereas all other European early farmers show greater genetic similarity to modern-day Sardinians. These genetic links suggest that Basques and their language may be linked with the spread of agriculture during the Neolithic. Furthermore, all modern-day Iberian groups except the Basques display distinct admixture with Caucasus/Central Asian and North African groups, possibly related to historical migration events. The El Portalón genomes uncover important pieces of the demographic history of Iberia and Europe and reveal how prehistoric groups relate to modern-day people.

It must be said to begin with that the Atapuerca samples are actually similarly related to Basques as to Sardinians: they have more Paleoeuropean admixture than Sardinians and early European farmers but not quite as much as Basques. The various formal analyses, such as the one displayed to the right (fig. 3-B) confirm this intermediate position between what I'd call First Neolithic and Atlantic Neolithic genetic configurations, which can be directly associated to modern Sardinians and modern Basques respectively.
Other data such as the mtDNA pool or lack of the lactase persistance allele also places them rather in the
First Neolithic group in spite of their greater Paleoeuropean admixture, which is undeniable. The El Portalón samples have much lower frequencies of mtDNA H and U than
Neolithic Basques or
Burgundians, let alone the "hyper-modern" Neolithic
Portuguese with their >80% mtDNA H (Chandler et al. 2005).
Similarly their lack of the T-13910 lactase persistence allele, dominant among modern Basques and many other Western Europeans, and
already detected in at least some Chalcolithic Basques from an intermediate area (Upper Ebro banks), suggest that the old archaeological and anthropometric narrative of Mediterranean colonists migrating up the Ebro and establishing intermediate (but still rather Mediterranean) populations in the Upper Ebro banks, somewhat distinct of proto-Basques proper of a distinctive Pyrenean (Keltid?) type, was not completely wrong. It is true that the Pyrenean type cannot be anymore considered a pure derivate from Paleoeuropeans but rather a mixed population with strong Mediterranean Neolithic input but there is still some distinction very apparent in the archaeogenetics we know so far that cannot be totally ignored.
Not a highlight of the study or the press release but I think it is very worth mentioning that the Bronze Age ATP9 woman also shows strong affinity with Britons, particularly Cornish and Scots. In general there is stronger mainland European affinity for this sample but still Basques and Sardinians, as well as the mentioned Britons, are the closest matches.
The general details of the samples are as follow:
As you can see only four samples had sufficiently good coverage to be considered for most analysis. Mitochondrial DNA, as usual, is the exception (all are good enough) but Y-DNA cannot be reliably assailed from such a poor quality sequence, nor actually much more that is not fuzzy.
Stopping for a moment on Y-DNA, it must be said that haplogroup H2 was formerly known as F3, being a rare West Eurasian haplogroup (with some presence in the Persian Gulf and Zagros mountains area, as well as a scatter through Europe) which has seen its phylogeny recently refined under H (otherwise a South Asian and Roma lineage).
I2a2a is not the typical Sardinian and Pyrenean lineage that is generally considered to be part of the Cardium Pottery package, originating probably in the Balcans, even I first thought it would be (thanks to Krefter for the correction). Instead it is some other Paleoeuropean lineage, which is today most concentrated in Northern Europe (→
map).
Genetiker claims that ATP3 should be R1b1a2-M269, while ATP17 would be also I2a2a. However given the very low coverage of these genomes, I would take such claims with great caution. As I've written somewhere else the question is not anyhow if there are some R1b of any sort here or there because the M269 → L11 stage has left only a scattered legacy, except for two large subhaplogroups: S116/P312 and U106. These two subhaplogroups are the big mystery and for them we only have so far late Chalcolithic
terminus ante quem dates from Bell Beaker Germany and Corded Ware Sweden respectively. The lack of Atlantic Neolithic samples, be them British, French, Basque or Portuguese surely has a lot to do with this lack of evidence because, you know, follow the trail of
modernity traits such as early "modern" mtDNA pools (Neolithic Basques and North French, Portuguese also) or early presence of the lactose tolerance allele (Chalcolithic Basques and Swedes). The answer to this pressing question must be in the Atlantic basin of Europe, just sequence it for Chaos' sake!
As for the mtDNA, the genetic pool is partly typical Neolithic (K, J, X) but with notable highlights: on one side, there are 3/8 U5 lineages, which are surely a legacy of the admixture with Paleoeuropeans, and, on the other side there is this H3 haplogroup that is nowadays important especially in Southwestern Europe. Unlike H1 or H6, which are known to have been carried by Paleoeuropeans, H3 so far has only been found among Neo-Europeans (i.e. from Neolithic onwards) but we do not know so far its ultimate origins for sure, as happens with haplogroup V (similar situation).
In any case the low frequency of haplogroup H, makes the overall pool not yet "modern", unlike what happens with at least some Neolithic Basques (Paternabidea) and certainly with proto-Basques from Chalcolithic onwards, as well as other mentioned groups like North French Neolithic, Portuguese Neolithic and later also German Bell Beaker peoples. A more complete comparison of various ancient mtDNA pools can be found in fig. S4.
Going back to the main focus of the study, which is autosomal DNA, I guess that we can continue with the following excerpt from the ADMIXTURE analysis:
 |
| Fig. 3(A) - Population structure of ancient and modern-day individuals. (A) Admixture fractions among modern-day individuals from Eurasia and North Africa together with 16 ancient individuals. Only ancient and modern-day individuals from Southwestern Europe are shown (see Dataset S1 for the complete plot with all individuals). Admixture components are labeled based on the populations/geographic regions in which they are modal. |
The two main components are (1) the Paleoeuropean of HG modal, whose fractions may well be close to real in this case, at least judging on the strict alignment of Europeans between Paleoeuropeans and West Asians provided by the PCA with North African samples (see below) and (2) the West Asian or EEF modal. We see that the First Neolithic populations were around 20-25% Paleoeuropean (the orange fraction would be West Asian or mostly so), while the Atapuerca samples show double HG scores, c. 40-50%. Modern Basques are even higher, around 50-60% Paleoeuropean in this analysis (although obviously a small fraction should be attributed to the Early Farmers' inflow).
In any case we see that in general the increase of the Paleoeuropean fraction is very notable in Atapuerca and Gokhem, and was plausibly even greater among other Atlantic early farmers, judging on the modern Basque and Gascon ("French South") data.
The next most relevant component is the Caucasus/Central Asian one, which should be attributed to the Indoeuropean expansion almost certainly. These would also have contributed with a proportional fraction of Paleoeuropean blood (maybe double than the black segment or something in that line). We see that this Indoeuropean influence is most important among the French and much less relevant in Iberia (but still much greater than among Basques or Sardinians). Nothing really new in this after Lazaridis 2014, Haak 2015 and Alentoft 2015, just a complementary perspective on the same issue.
As for the North African component it seems to follow the same pattern of it being mostly a feature of the Western third of the Iberian Peninsula rather than something attributable to "historical events" (such as the Muslim period or the Phoenician conquest, much more influential in the South and East instead). Again lack of ancient DNA from those regions have hindered the understanding of the origin of this component which can be either (1) a Neolithic founder effect or (2) a Paleolithic founder effect dating from as early as the Solutrean-Oranian interaction around the Last Glacial Maximum. The fact that La Braña in some analysis shows North African or otherwise African affinities suggests that it is a Paleolithic element that has remained basically a Western Iberian thing (excepted the occasional founder effect such as certain district of Northern Wales and some diffuse scatter of related markers like E1b-M81 along the Atlantic coast of Europe).
If this interpretation is correct, then it strongly weights against the hypothesis of widespread (Western) Iberian origin of Chalcolithic (Megalithic, Bell Beaker) founder effects through Western Europe, because we would see it everywhere. Instead I'm much more inclined for a major role of what are now the Western parts of France, which were clearly involved in the (late) Neolithic colonization of Britain. I'd rather advocate for a "French" origin of Y-DNA haplogroup R1b-S116 for example.
True that studying the Hexagon is sadly hampered by a hostile bureaucracy, legal framework and state ideology but no such obstacle seems to impede the study of ancient Britons, for example. In due time I guess.
Another highlight is the TreeMix graph. However I must say that I am much more comfortable with the S10 tree rather than the highlighted one in the main body of the study. The main difference is that Ötzi (Iceman) is in this one placed near Gokhem and also with some Pitted Ware type admixture. And this is new and therefore a bit disturbing. Instead the S10 tree is not so surprising:
Compare with the simplified 2-A tree to the right...
You choose, as not enough data is provided in the paper to argument one choice above the other.
It is apparent in any case that Gokhem seems to have strong admixture from a source that is precursor of Pitted Ware Ajv58. This may also be the case of Ötzi (?), although it'd be weaker.
Also, in spite of claims of La Braña admixture in Atapuerca I do not see that element clear in either graph. In one it is apparent that the Paleoeuropean influence is rather Lochsbour-like, while in the other it seems something rather ancestral to both WHG individuals.
Confused? Well, what about the pre-Motala 46% admixture in Mal'ta 1? That is indeed new and it is consistent in both graphs. I would think it is suggesting that, rather than Ma1 (ANE) admixture in SHG, what we have is an ancient flow from Europe to LGM Siberia. Which can that be? Gravettian culture only. Oddly enough it makes some sense but, if this is correct, then it should also say that all the ANE buzz was a bit nonsensical after all. Confounding factors at play.
To finish this review I'll copy here the various Principal Component Analyses, eye-candy, which in this case have diverse sampling strategies. Oddly enough for someone who has asked for Europe-only PCAs, it seems to me that in this case the one including West Asians is a better approximation to the reality of Atapuerca's ancients:
 |
| Fig S7 |
As you can see, the three Chalcolithic samples properly sit in this case right between Basques and Sardinians. The position of the Bronze Age sample should be explained by the British affinity, rather than being truly closer to Spaniards.
All this is less clear in the Europe-only analysis:
 |
| Fig 2-C |
They do appear correctly between Basques and Sardinians but a shallow look could make them look as almost Spaniards, something that is not quite correct. Again the Bronze Age sample is pulled towards Britain, incidentally (and misleadingly) overlapping Spaniards.
Someone somewhere asked for a PCA with North African samples. Well, this study also includes one. However no North African influence is apparent in any European, ancient or modern, so the best use we can have for it is using the lineal reorganization of Europeans between the Paleoeuropean and the West Asian polarities as a ruler of sorts to estimate the levels of admixture of the various populations:
 |
| Fig S6 |
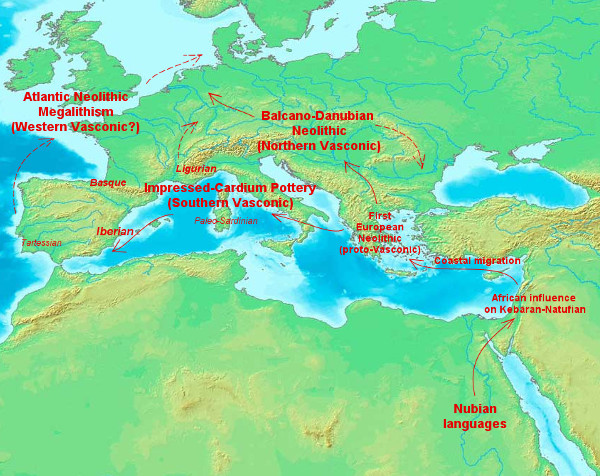
And that's all for today. Soon to come (hopefully): new Y-DNA age estimates (I promised more than a month ago, shame on me), genetics of Baltic peoples (new study) and the Vasco-Nubian linguistic connection (something I've been ruminating for more than a year now).
Update (Oct 16): I just stumbled on
this review[es] of a Bronze Age weapon hoard in Los Cascajos (Upper Rioja, unrelated to the Neolithic site of the same name in Navarre), not too far from Atapuerca, and the authors mention a practical identity in design between one of the swords and another one from Stratford (England), which is known to be of local manufacture. Hence he ponders that at least some of the weapons could have been imported from Great Britain.
The, very apparently ritual, destruction of the weapons, including the use of fire, before their burial also resembles a lot British rituals of the same period.
All this, along with other well known Iberia-Britain exchanges in the Bronze Age, may help to support the notion of ATP9 being partly of British ancestry.














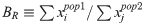







{kind=link}
{kind=link}